The protocol presented here describes an intravital microscopy-based approach to visualize intestinal epithelial leakage and observe cell shedding performance in the gut in real-time. Briefly, mice are anesthetized and submitted to surgical preparation in order to expose the surface of the small intestine mucosa. IECs are then stained via topical application of acriflavine; while luminal rhodamine B-dextran is used as tracers to detect transmucosal passage from the lumen to the sub-epithelial space. Thus, the surgical preparation and the anesthetized mouse are placed on a slide mounted in a Petri dish and imaged over time using CLSM (Figure 1). Post-acquisition analysis permits the calculation of epithelial cell shedding rate (number of cell shedding events/minute/length of basal membrane) as well as the percentage of transitory leakage (paracellular permeability) and "permeable cells" (transcellular permeability) at a determined time point.
In order to induce alterations of epithelial integrity, we took advantage of the previously described IEC-specific GGTase-deficient conditional mouse model (Pggt1biΔIEC mice), generated via the LoxP-Cre system24. As described before, Pggt1biΔIEC mice (Figure 2A) developed severe intestinal pathology as shown by increased histological damage score in small intestine (Figure 2B). Increased intestinal epithelial permeability could be detected via tracer in vivo experiments using orally administered FITC-dextran (4 kDa) (Figure 2C), and then confirmed via intravital microscopy (Figure 2D-2E). While rhodamine dextran is restricted to the luminal compartment in control mice, we could detect the tracer within the sub-epithelial compartment upon abrogation of GGTase expression within IECs in Pggt1biΔIEC mice. During image acquisition, cell shedding events could be identified as cells moving out of the epithelial monolayer into the lumen, leading to temporary gaps in the sealing of the epithelium, which are finally closed by the contact between neighboring cells, so called zip-effect (Figure 3A). We could clearly observe these gaps, what we call temporary epithelial leakage both in control and Pggt1biΔIEC mice, although the frequency of these phenomenon was higher in the latter (Figure 3B,3C). Interestingly, we could also identify other cells where dextran could be detected intracellularly, so called "permeable cells"; these events occurred mainly in Pggt1biΔIEC mice (Figure 3B,3D). Together, taking advantage of the here presented intravital microscopy approach, we could determine that impaired epithelial integrity in Pggt1biΔIEC mice leads to cell shedding performance alterations and increased para- and trans-cellular epithelial permeability in the gut.
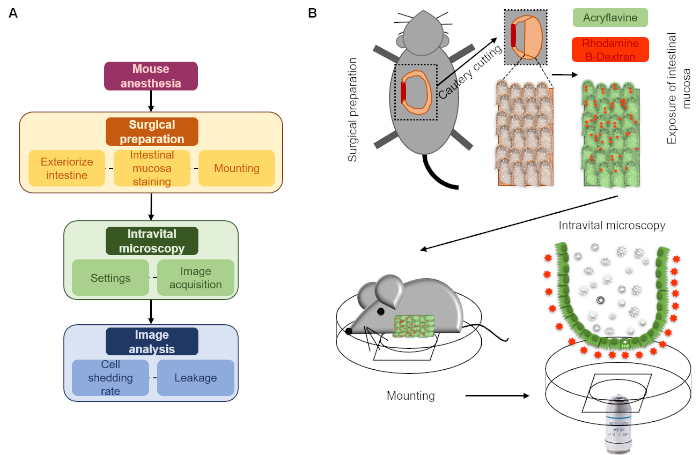
Figure 1: Schematic description of the here presented intravital microscopy approach. (A) Flow chart. (B) Diagram. After surgical preparation, intestinal mucosa topically stained with acriflavine and rhodamine dextran is mounted on a cover-slide embedded on a Petri dish to allow perfusion with a saline solution (luminal surface down). The intestinal mucosa is then imaged using a CLSM microscope over time. Please click here to view a larger version of this figure.

Figure 2: Impaired epithelial integrity in Pggt1biΔIEC mice leading to increased intestinal permeability. (A) Western blot showing tamoxifen-induced abolished GGTase-1B expression within IECs in Pggt1biΔIEC versus control mice. (B) Histological score of the small intestine from the control and Pggt1biΔIEC mice. (C) Quantification of intestinal epithelial permeability in vivo measured by transmucosal passage of orally administered FITC-Dextran (4 kDa). (D) Diagram describing the direction of image acquisition, from the lumen downwards to the villus axis. Representative pictures from a z-stack. (E) Representative pictures of intravital microscopy using topically applied acriflavine and rhodamine B-dextran from control and Pggt1biΔIEC mice, as described in this manuscript. Data are expressed as Mean ± SEM. Non-paired t-test. * P value ≤ 0.05; *** P value ≤ 0.0001. Please click here to view a larger version of this figure.
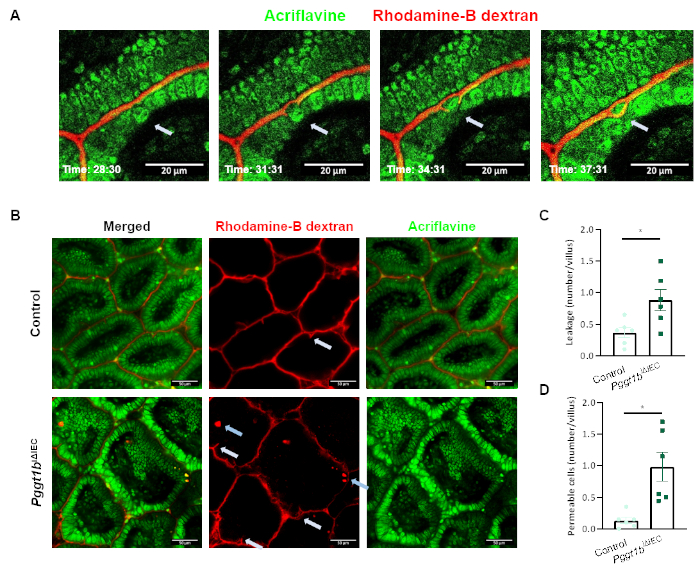
Figure 3: Epithelial cell shedding performance and para/trans-cellular epithelial permeability using intravital microscopy. (A) Representative pictures showing a cell shedding event at real-time (white arrow). The shed cell is extruded from the epithelial monolayer to the lumen. Neighboring cells seal the temporary leakage (zip-effect) to avoid loss of barrier function. (B) Representative picture showing leakage (white arrows) and permeable intestinal epithelial cells (blue arrows). (C-D) Quantification of temporary leakage (C) and permeable cells (D) in control and Pggt1biΔIEC mice. Data are expressed as Mean ± SEM. Non-paired t-test. * P value ≤ 0.05; *** P value ≤ 0.0001.Please click here to view a larger version of this figure.
