The following results were obtained when a collection of antibiotic-producing strains of interest were dereplicated using the ARP and/or MARP.
A diagram of the ARP/MARP dereplication workflow is depicted in Figure 1, and library plate maps are shown in Supplemental Figure 1 and Supplemental Figure 2. Figure 2 demonstrates a positive dereplication result wherein the environmental extract WAC 8921 is identified as a chloramphenicol producer. Figure 3 shows a lack of ARP growth entirely, which indicates the presence of either an unknown antibiotic or a less commonly found antibiotic that is not accounted for in the ARP/MARP library plate. Figure 4 demonstrates a growth pattern that is unique to the MARP because of its utilization of both wildtype E. coli BW25113 and a hyperpermeable and efflux deficient mutant E. coli BW25113 ΔbamBΔtolC. This result suggests the presence of a compound with antimicrobial activity that is unable to surpass an intact outer membrane. Figure 5 shows an E. coli growth pattern that suggests the improper sterilization of pinning tools and Figure 6 shows an example of ARP/MARP frozen stock library plate contamination. Figure 7 demonstrates what happens if the agar overlay is pierced during dereplication. Lastly, Figure 8 shows MHB overlay related contamination that can occur during the dereplication process.
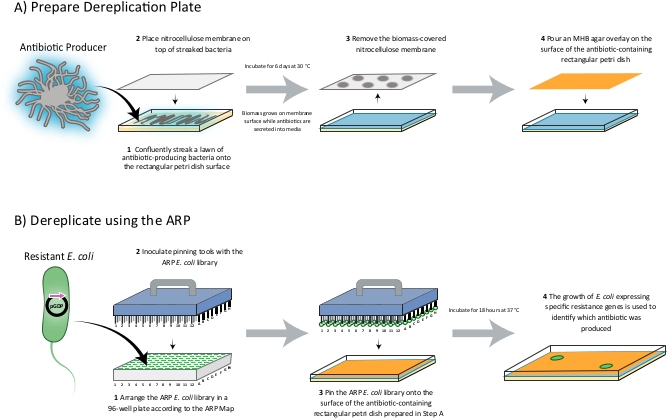
Figure 1: A schematic of the dereplication process. The producing strain to be dereplicated is streaked onto a rectangular Petri dish as a lawn and a nitrocellulose membrane is placed on top. The plate is then incubated for 6 days wherein the producing-strain related biomass grows on the surface of the membrane while and secondary metabolites produced are secreted into the Petri dish media. After a 6-day fermentation period, the membrane is removed and an MHB overlay is added to the surface of the antibiotic-containing media to provide a smooth surface for pinning. The ARP/MARP E. coli library, which is arranged in a 96-well plate format according to the ARP/MARP Maps, is then pinned onto the surface of the overlay. After incubating the tray overnight at 37 °C, the growth of E. coli strains expressing specific resistance genes indicates the identity of the compound produced. Please click here to view a larger version of this figure.

Figure 2: Dereplication of a known antibiotic. The producing strain WAC 8921 was dereplicated using the ARP template. Growth of E. coli BW25113 ΔbamBΔtolC pGDP1:CAT on the surface of the MHB agar overlay indicates that WAC 8921 is a chloramphenicol producer. Please click here to view a larger version of this figure.

Figure 3: Dereplication of an unknown antibiotic. The producing strain WAC 9941 was dereplicated using the ARP template. A lack of E. coli library growth was seen on the surface of the rectangular Petri dish, indicating that either WAC 9441 is producing an unknown antimicrobial compound or a rare antibiotic that is not accounted for in the ARP. Please click here to view a larger version of this figure.

Figure 4: Identification of an antimicrobial compound that cannot traverse an intact outer membrane. The producing strain WAC 4178 was dereplicated using the MARP template. Strains of E. coli BW25113 are capable of growing on the surface of the secondary metabolite-containing media, whereas all strains of E. coli BW25113 ΔbamBΔtolC cannot grow. This suggests that WAC 4178 is producing an antimicrobial compound that cannot traverse an intact outer membrane. Please click here to view a larger version of this figure.

Figure 5: Contamination due to non-sterile pinning tools. The producing strain WAC 7094 was dereplicated using the ARP template. The presence of E. coli library growth in areas that did not have an assigned E. coli strain suggests that the pinning tools used to inoculate the MHB agar overlay of the rectangular Petri dish were not properly sterilized. This results in the transfer of unknown E. coli strains across the overlay. Please click here to view a larger version of this figure.

Figure 6: Contamination due to a contaminated frozen stock ARP/MARP template. The producing strain WAC 3683 was dereplicated using the ARP template. Three distinct E. coli colonies grew on the rectangular Petri dish surface: two correspond with E. coli BW25113 ΔbamBΔtolC expressing STAT, a streptothricin resistance enzyme, and the other corresponds to E. coli BW25113 ΔbamBΔtolC expressing VIM-2ss, a β-lactam resistance enzyme. Due to a lack of replicating blaVIM2ss colony growth, in addition to the lack of cross-resistance known to occur between these two antibiotic classes, it can be assumed that a strain other than E. coli ΔbamBΔtolC pGDP1: blaVIM2ss is growing in the respective frozen library plate well. Please click here to view a larger version of this figure.

Figure 7: Pierced MHB agar overlay. The producing strain WAC 5106 was dereplicated using the ARP template. This strain was found to be a streptomycin producer, as indicated by the growth of E. coli BW25113 ΔbamBΔtolC pGDP3:aph(6)-Ia. Puncture holes can be seen on the surface of the MHB agar overlay along the perimeter of the plate. While this does not affect the dereplication results, it can make the data difficult to interpret at first glance. Please click here to view a larger version of this figure.

Figure 8: Contamination of the MHB agar overlay. Contaminated MHB agar produces an irregular growth pattern on the surface of the overlay that becomes visible after incubating the plate overnight at 37 °C. Although E. coli growth may still be visible through the contamination, it is advised to repeat the experiment before extrapolating data from the plate. Please click here to view a larger version of this figure.
| Plasmid | Selectable Marker |
| pGDP1 | Kanamycin 50 μg/mL |
| pGDP2 | |
| pGDP3 | Ampicillin 100 μg/mL |
| pGDP4 | |
| None | – |
Table 1: Selectable markers used in the pGDP plasmid series. Streak the ARP/MARP E. coli strains onto LB agar Petri dishes containing the appropriate selectable marker at the right concentration for each plasmid.
| Media | Ingredient | Amount |
| SAM | Glucose | 15 g |
| Soya peptone | 15 g | |
| NaCl | 5 g | |
| Yeast extract | 1 g | |
| CaCO3 | 1 g | |
| Glycerol | 2.5 mL | |
| ddH2O | To 1 L | |
| Bennett's | Potato starch | 10 g |
| Casamino acids | 2 g | |
| Yeast extract | 1.8 g | |
| Czapek mineral mix | 2 mL | |
| Agar (optional) | 15 g | |
| ddH2O | To 1 L | |
| Czapek mineral mix | KCl | 10 g |
| MgSO4∙7H2O | 10 g | |
| NaNO3 | 12 g | |
| FeSO4∙7H2O | 0.2 g | |
| Concentrated HCl | 200 µL | |
| ddH2O | To 100 mL |
Table 2: Recipes for SAM and Bennett’s media, and Czapek mineral mix. Adjust SAM and Bennett’s to pH 6.8 before autoclaving, and filter sterilize the Czapek mineral mix.
| Antibiotic Class | Antibiotic | Resistance Gene | E. coli Strain | Well Position |
| Aminoglycosides | Streptomycin | aph(3’’)-Ia | ΔbamBΔtolC BW25113 | B3, G10 |
| 2- Deoxystreptamine | rmtB | ΔbamBΔtolC BW25113 | F3, C10 | |
| Apramycin | apmA | ΔbamBΔtolC BW25113 | C5, F8 | |
| Spectinomycin | aph(9)-Ia | ΔbamBΔtolC BW25113 | B5, G8 | |
| β-lactams | Penicillin | NDM-1 | ΔbamBΔtolC BW25113 | B4, G9 |
| Cephalosporin | ||||
| Carbapenam | ||||
| Lincosamides | Lincosamides | ermC | ΔbamBΔtolC BW25113 | D4, E9 |
| Macrolides | Macrolides | ermC | ||
| Type B Streptogramins | Type B Streptogramins | ermC | ||
| Type A Streptogramins | Type A Streptogramins | vatD | ΔbamBΔtolC BW25113 | C3, F10 |
| Streptothricin | Streptothricin | STAT | ΔbamBΔtolC BW25113 | D3, E10 |
| Tetracyclines | Tetracycline | tet(A) | ΔbamBΔtolC BW25113 | D5, E8 |
| Chloramphenicols | Chloramphenicols | CAT | ΔbamBΔtolC BW25113 | E4, D9 |
| Fosfomycins | Fosfomycins | fosA | ΔbamBΔtolC BW25113 | F6, C7 |
| Rifamycins | Rifamycins | arr | ΔbamBΔtolC BW25113 | E3, D10 |
| Polymyxins | Polymyxins | MCR-1 | wild-type BW25113 | C6, F7 |
| Echinomycins | Echinomycins | uvrA | ΔbamBΔtolC BW25113 | F4, C9 |
| Sideromycins | Albomycin | fhuB mutant | ΔbamBΔtolC BW25113 | C4, F9 |
| Tuberactinomycins | Viomycin | vph | ΔbamBΔtolC BW25113 | F5, C8 |
| N/A | N/A | N/A | wild-type BW25113 | C1, C12, F1, F12, E5, D8 |
| N/A | N/A | N/A | ΔbamBΔtolC BW25113 | A1, A12, B1, B12, D6, D7, E6, E7, G1, G12, H1, H12 |
Table 3: Well designation table for the minimal ARP strains. This table indicates which well of a 96-well plate that each of the minimal ARP strains can be found in according to the minimal ARP library plate map. The table also lists which antibiotic class each gene confers resistance to. Please note that some genes may confer resistance to more than one antibiotic within the given antibiotic class.
| Antibiotic Class | Antibiotic | Resistance Gene | E. coli Strain | Well Position |
| Aminoglycosides | Streptomycin | aph(3’’)-Ia | ΔbamBΔtolC BW25113 | B2, G11 |
| aph(6)-Ia | ΔbamBΔtolC BW25113 | C6,F7 | ||
| Spectinomycin | aph(9)-Ia | ΔbamBΔtolC BW25113 | A2, H11 | |
| Gentamicin | aac(3)-Ia | ΔbamBΔtolC BW25113 | A3,H10 | |
| ant(2’’)-Ia | ΔbamBΔtolC BW25113 | A5, H8 | ||
| aph(2’’)-Id | ΔbamBΔtolC BW25113 | A4, H9 | ||
| armA | ΔbamBΔtolC BW25113 | A6, H7 | ||
| aac(6’)-aph(2’’)-Ia | ΔbamBΔtolC BW25113 | B5,G8 | ||
| Kanamycin | aph(3’)-Ia | ΔbamBΔtolC BW25113 | B4, G9 | |
| aph(3’)-Illa | ΔbamBΔtolC BW25113 | B3, G10 | ||
| Hygromycin | aph(4)-Ia | ΔbamBΔtolC BW25113 | B6,G7 | |
| β-lactams | Amoxicillin | TEM-1 | ΔbamBΔtolC BW25113 | F6, C7 |
| Ceftazidime | CTX-M-15 | ΔbamBΔtolC BW25113 | F5, C8 | |
| Oxacillin | OXA-10 | ΔbamBΔtolC BW25113 | G5, B8 | |
| OXA-48 | ΔbamBΔtolC BW25113 | H5, A8 | ||
| Meropenem | IMP-7ss | ΔbamBΔtolC BW25113 | G4, B9 | |
| KPC-2 | ΔbamBΔtolC BW25113 | G6, B7 | ||
| NDM-1 | ΔbamBΔtolC BW25113 | H6, A7 | ||
| Imipenem | VIM-2 | ΔbamBΔtolC BW25113 | F4, C9 | |
| Lincosamides | Lincosamides | ermC | ΔbamBΔtolC BW25113 | C4, F9 |
| lnu(A) | ΔbamBΔtolC BW25113 | C5, F8 | ||
| Macrolides | Macrolides | ermC | ΔbamBΔtolC BW25113 | C4, F9 |
| mphA | ΔbamBΔtolC BW25113 | C3, F10 | ||
| mphB | ΔbamBΔtolC BW25113 | C2, F11 | ||
| Type B Streptogramins | Type B Streptogramins | ermC | ΔbamBΔtolC BW25113 | C4, F9 |
| Vgb | ΔbamBΔtolC BW25113 | D4, E9 | ||
| Type A Streptogramins | Type A Streptogramins | vatD | ΔbamBΔtolC BW25113 | D5, E8 |
| Streptothricin | Streptothricin | STAT | ΔbamBΔtolC BW25113 | D3, E10 |
| Tetracyclines | Tetracycline | tet(M) | ΔbamBΔtolC BW25113 | E5, D8 |
| Chloramphenicols | Chloramphenicols | CAT | ΔbamBΔtolC BW25113 | E4, D9 |
| Fosfomycins | Fosfomycins | fosA | ΔbamBΔtolC BW25113 | H4, A9 |
| Rifamycins | Rifamycins | arr | ΔbamBΔtolC BW25113 | E3, D10 |
| N/A | N/A | N/A | ΔbamBΔtolC BW25113 | A1, A12, B1, B12, D6, D7, E6, E7, G1, G12, H1, H12 |
Table 4: Well designation table for the ARP strains. This table indicates which well of a 96-well plate that each of the ARP strains can be found in according to the ARP library plate map. The table also lists which antibiotic class each gene confers resistance to. Please note that some genes may confer resistance to more than one antibiotic within the given antibiotic class.
Supplemental Figure 1: Library plate map used for the original antibiotic resistance platform (ARP) template. Organize the respective E. coli strains in a 96-well plate using this format to ensure that all necessary controls and duplicates are included. This figure has been modified from Cox et al.10. Please click here to download this figure.
Supplemental Figure 2: Library plate map used for the minimal antibiotic resistance platform (MARP) template. Organize the respective E. coli strains in a 96-well plate using this format to ensure that all necessary controls and duplicates are included. Please click here to download this figure.
