Figure 1 shows the gating strategy used to divide the TCM or TEM of CD8+ or CD4+ T cells from a representative JEV stimulation group of JE-vaccinated children. The FSC-A/SSC-A dot plot is used to identify lymphocytes, and the FSC-A/FSC-W dot plot is used to identify single cells. Viable cells are selected on the live/dead/SSC-A dot plot. The CD3/SSC-A dot plot is used to identify the CD3+ T cells. The CD4/CD8 dot plot is used to identify the CD3+ CD4+ and CD3+ CD8+ T cells, respectively. The TCM (CD27+ CD45RO+) and TEM (CD27– CD45RO+) of CD8+ or CD4+ T cells are selected separately on the CD45RO/CD27 dot plot. Cells with the phenotype of CD107a+, IL-2+, TNF-α+, IFN-γ+, and MIP-1α+ from CD8+ TCM (Figure 1F), CD8+ TEM (Figure 1G), CD4+ TCM (Figure 1H), and CD4+ TEM (Figure 1I) are selected separately by drawing the corresponding rectangular gate.
The polyfunctional characterization of central and effector memory CD4+ and CD8+ T cell responses (including degranulation, cytokines, and chemokines) to JEV in JE-vaccinated and unvaccinated children are shown in Table 3. These results indicate that increased levels of CD107a, IFN-γ, TNF-α, and IL-2 were detected in the CD8+ TCM cells of the vaccinated children after JEV stimulation compared to those in unvaccinated children. The level of MIP-1α did not differ between the two groups. TEM cells are thought to exert antiviral effects directly upon restimulation with JEV. Higher levels of CD107a and IFN-γ were detected in the CD8+ TEM cells of the vaccinated group under JEV stimulation compared with the unvaccinated group. However, TNF-α, IL-2, and MIP-1α were not significantly elevated in those positive cells.
The JEV antigen successfully induced higher levels of CD107a, IFN-γ, TNF-α, and MIP-1α in the CD4+ TCM cells of the vaccinated group compared with the unvaccinated group. However, the proportion of IL-2+ cells did not differ between the two groups under JEV stimulation. The proportion of the CD107a+, IFN-γ+, and TNF-α+ subsets of CD4+ TEM cells in the vaccinated group was higher in the presence of JEV than in the unvaccinated group.
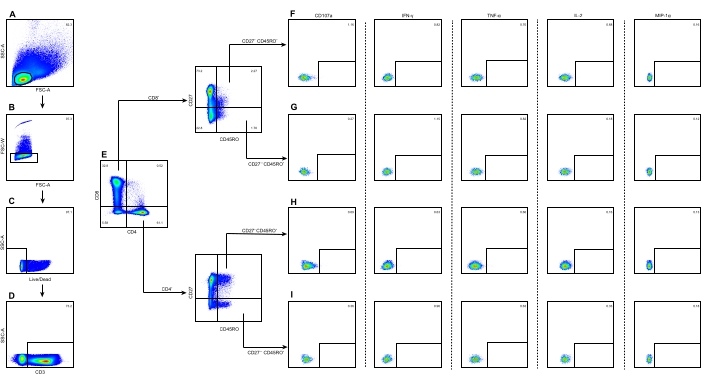
Figure 1: Illustration of the gating strategy to identify the TCM or TEM of CD8+ or CD4+ T cells and their subsets. (A) Lymphocytes were identified using the FSC-A/SSC-A dot plot. (B) Single cells were identified using the FSC-A/FSC-W dot plot. (C) Live cells were identified using the live/dead/SSC-A dot plot. (D) CD3+ T cells were identified using the CD3/SSC-A dot plot. (E) CD3+ CD4+ T and CD3+ CD8+ T cells were identified using the CD4/CD8 dot plot. (F) CD8+ TCM with the phenotype of CD107a+, IL-2+, TNF-α+, IFN-γ+, and MIP-1α+were subdivided separately. (G) CD8+ TEM with the five phenotypes were subdivided separately. (H) CD4+ TCM with the five phenotypes were subdivided separately. (I) CD4+ TEM with the five phenotypes were subdivided separately. The numbers on each panel represent the percentage of cells in the gates. The color coding represents the frequency of occurrence of the cells, where red is the highest frequency and blue is the lowest. Abbreviations: SSC-A = side scatter-area; FSC-A = forward scatter-area; FSC-W = forward scatter-width. Please click here to view a larger version of this figure.
| Antibody Target | Conjugated Fluorophore | Dosage | Clone | Isotype | Excitation laser line | Ex-Max (nm) | Em-Max (nm) |
| CD3 | BV650 | 2 µL | SK7 | Mouse IgG1, κ | Violet | 407 | 650 |
| CD4 | BUV395 | 2 µL | SK3 | Mouse IgG1, κ | UV | 348 | 39 |
| CD8 | BV421 | 2 µL | SK1 | Mouse IgG1, κ | Violet | 407 | 421 |
| CD27 | BUV737 | 2 µL | L128 | Mouse IgG1, κ | UV | 350 | 737 |
| CD45RO | BV480 | 2 µL | UCHL1 | Mouse IgG2a, κ | Violet | 436 | 478 |
| CCR7 | BUV737 | 2 µL | 3D12 | Rat IgG2a, κ | UV | 350 | 737 |
| CD45RA | BV480 | 2 µL | HI100 | Mouse IgG2b, κ | Violet | 436 | 478 |
| CD107a | BV605 | 2 µL | H4A3 | Mouse IgG1, κ | Violet | 407 | 602 |
| IL-2 | BV785 | 2 µL | MQ1-17H12 | Rat IgG2a, κ | Violet | 407 | 786 |
| IFN-γ | FITC | 2 µL | 4S.B3 | Mouse IgG1, κ | Blue | 494 | 520 |
| TNF-α | PE | 2 µL | MAb11 | Mouse IgG1, κ | Yellow-Green | 496, 564 | 578 |
| MIP-1α | APC | 2 µL | 11A3 | Mouse IgG2a, κ | Red | 650 | 660 |
| Zombie NIR | APC-Cy7 | 1 µL | – | – | Red | 650 | 779 |
Table 1: The color fluorescent antibody staining protocol for flow cytometric analyses. Abbreviations: BV = brilliant violet; BUV = brilliant ultraviolet; FITC = fluorescein isothiocyanate; PE = phycoerythrin; APC = allophycocyanin.
| Compensation reference of flow cytometry (%) | ||||||||||||
| FITC | APC | APC-Cy7 | BV421 | BV480 | BV605 | BV560 | BV785 | BUV395 | BUV737 | PE | ||
| FITC | 100 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| APC | 0 | 100 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| APC-Cy7 | 0 | 0 | 100 | 0 | 0 | 0 | 0 | 65 | 0 | 44 | 0 | |
| BV421 | 0 | 0.6 | 0 | 100 | 10.5 | 3 | 15 | 3 | 0.4 | 0 | 0 | |
| BV480 | 3 | 0 | 0 | 46 | 100 | 19 | 0 | 6.1 | 0 | 0 | 0 | |
| BV605 | 0 | 0 | 0 | 3.6 | 0 | 100 | 94 | 10 | 0 | 9 | 14 | |
| BV560 | 0 | 5.5001 | 0 | 2 | 1 | 7.1 | 100 | 9 | 0 | 10 | 0 | |
| BV785 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 100 | 0 | 30 | 0 | |
| BUV395 | 0 | 0 | 0 | 1.7 | 0 | 0 | 0 | 0 | 100 | 0 | 0 | |
| BUV737 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 3.7001 | 3 | 100 | 0 | |
| PE | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 100 | |
Table 2: The parameters of compensation for flow cytometric analyses. Abbreviations: BV = brilliant violet; BUV = brilliant ultraviolet; FITC = fluorescein isothiocyanate; PE = phycoerythrin; APC = allophycocyanin.
| Group of TPF | Polyfunctional characterization (%) | |||||||
| Stimulus | CD107a | IFN-γ | TNF-α | IL-2 | MIP-1α | |||
| CD8+ T cells | TCM | Vaccinated | JEV | 1.50±0.48 | 1.60±0.69 | 0.66±0.32 | 0.54±0.27 | 0.16±0.11 |
| Ctrl | 0.51±0.38 | 0.31±0.13 | 0.28±0.13 | 0.37±0.20 | 0.02±0.05 | |||
| Unvaccinated | JEV | 0.38±0.19 | 0.20±0.03 | 0.19±0.09 | 0.16±0.04 | 0.19±0.09 | ||
| Ctrl | 0.50±0.28 | 0.27±0.09 | 0.19±0.05 | 0.23±0.14 | 0.08±0.11 | |||
| P value* | – | 0.00 | 0.00 | 0.02 | 0.01 | 0.70 | ||
| TEM | Vaccinated | JEV | 1.47±0.58 | 1.21±0.22 | 0.49±0.36 | 0.33±0.31 | 0.24±0.17 | |
| Ctrl | 0.82±0.48 | 0.39±0.15 | 0.16±0.18 | 0.23±0.256 | 0.09±0.21 | |||
| Unvaccinated | JEV | 0.41±0.25 | 0.14±0.09 | 0.15±0.11 | 0.20±0.07 | 0.15±0.13 | ||
| Ctrl | 0.34±0.13 | 0.21±0.15 | 0.17±0.10 | 0.19±0.19 | 0.01±0.02 | |||
| P value* | – | 0.01 | 0.00 | 0.08 | 0.13 | 0.36 | ||
| CD4+ T cells | TCM | Vaccinated | JEV | 1.55±0.43 | 1.16±0.24 | 0.57±0.25 | 0.29±0.18 | 0.19±0.07 |
| Ctrl | 0.44±0.27 | 0.26±0.20 | 0.15±0.06 | 0.12±0.06 | 0.04±0.07 | |||
| Unvaccinated | JEV | 0.28±0.08 | 0.21±0.08 | 0.17±0.03 | 0.21±0.16 | 0.10±0.03 | ||
| Ctrl | 0.31±0.13 | 0.26±0.06 | 0.14±0.04 | 0.25±0.13 | 0.04±0.04 | |||
| P value* | – | 0.00 | 0.00 | 0.01 | 0.47 | 0.04 | ||
| TEM | Vaccinated | JEV | 1.54±0.58 | 1.33±0.15 | 0.89±0.25 | 0.37±0.22 | 0.21±0.05 | |
| Ctrl | 0.46±0.49 | 0.35±0.30 | 0.13±0.08 | 0.14±0.09 | 0.05±0.11 | |||
| Unvaccinated | JEV | 0.27±0.09 | 0.23±0.10 | 0.30±0.15 | 0.23±0.03 | 0.14±0.06 | ||
| Ctrl | 0.35±0.21 | 0.24±0.14 | 0.19±0.20 | 0.25±0.08 | 0.05±0.07 | |||
| P value* | – | 0.00 | 0.00 | 0.00 | 0.19 | 0.08 | ||
| *Mann–Whitney U-test, PBMCs from vaccinated individual stimulated by JEV vs.PBMCs from unvaccinated individual stimulated by JEV were showed only. | ||||||||
Table 3: Polyfunctional characterization of the TCM and TEM of CD4+ or CD8+ T cell responses to JEV. Vaccinated, n = 5; unvaccinated, n = 5. Results are expressed as mean ± SD. P < 0.05 indicated a significant difference. *PBMCs from vaccinated individuals stimulated by JEV versus PBMCs from unvaccinated individuals stimulated by JEV only are shown. Abbreviations: TPFs = polyfunctional T cells; IFN-γ = interferon-γ; TNF-α = tumor necrosis factor-α; IL-2 = interleukin-2; MIP-1α = macrophage inflammatory protein-1α; TCM = central memory T cells; TEM = effector memory T cells.
