Following the presented protocol, different retinal cells were successfully visualized and tracked in vivo using a combination of AAV-mediated gene delivery and CSLO. AAV2-hSyn-eGFP effectively transduced RGCs, resulting in robust eGFP expression throughout the retina, as confirmed by CSLO and colocalization with the RGC-specific marker, RNA binding protein with multiple splicing (RBPMS), specifically found in the ganglion cell layer (Figure 2 and Figure 3). The protocol also confirmed the ability to target Müller glia using AAV9-GfaABC1D-eGFP. CSLO visualization revealed widespread eGFP expression, specifically within the inner retinal layers, consistent with the known major distribution segment of Müller glia. The GFP positive signal also highly colocalized with the Müller glia marker glutamine synthetase (GS), further confirming the specificity of this approach (Figure 2E,F and Figure 3B). These findings reaffirmed the versatility of AAV-aided gene delivery for studying different retinal cell types. To confirm the ability to target Müller glia, AAV9-GfaABC1D-eGFP was used. CSLO imaging revealed widespread eGFP expression, specifically within the inner retinal layers, consistent with the known distribution of Müller glia (Figure 2E,F). This selective expression pattern was further validated by the high degree of colocalization observed between the GFP signal and the Müller glia marker glutamine synthetase (GS) (Figure 3B).
To quantify the specificity and efficiency of the viral targeting approach for both RGCs and Müller glia, a colocalization analysis was performed to compare GFP fluorescence with standard cell-type markers (Figure 3B,D). A total of 6 mouse eyes were analyzed. For RGCs targeted with AAV2-hSyn-GFP, transduction efficiency was 64% ± 3%, while transduction specificity was 68% ± 3%. For Müller glia targeted with AAV9-GfaABC1D-GFP, transduction efficiency was 58% ± 2%, and transduction specificity was 71% ± 2%. These results demonstrate that this AAV-mediated gene delivery approach enables selective labeling of distinct retinal cell types, highlighting its versatility for studying cellular and molecular processes in the retina.
To assess the suitability of CSLO for longitudinal monitoring of retinal cell perturbations, a transient ONC model was performed in AAV2-hSyn-eGFP-infected mice. A progressive decline in eGFP signal and RGC numbers was observed over two weeks (Figure 4). RGC survival was quantified with three metrics: CSLO fluorescence intensity, in vivo cell counts for CSLO images, and ex vivo RGC counts of RBPMS-positive cells. After image processing as described in step 4, at least 10 regions of interest (ROIs) were cropped from each retina (6 eyes total), ensuring each ROI encompassed the same retinal location and exhibited clear cell signals at each time point. CSLO fluorescence intensity was then quantified by measuring the mean intensity within each cropped ROI at each time point. In vivo, cell counts were obtained by manually counting individual GFP-positive RGCs within the same cropped ROI at each time point. Lastly, retinas were harvested for RBPMS immunostaining, and RBPMS-positive cells were counted within 24 defined areas imaged per retinal sector for each retina. Significant negative correlations were found between all three RGC survival metrics (CSLO fluorescence intensity, in vivo cell counts, and ex vivo RBPMS-positive cell counts) and the time post-ONC (p < 0.001 for all, Figure 4C), demonstrating progressive RGC loss and degeneration. Importantly, no significant differences were observed across the different experimental groups in terms of initial RGC density or rate of decline, indicating a gross consistency in the ONC model and imaging approach. These findings are consistent with previous studies on RGC damage after ONC and validate the effectiveness of this CSLO-based protocol as a non-invasive and reliable tool for monitoring RGC health.
To further investigate RGC axonal projections within the visual pathway, a dual-labeling approach was used with the combination of AAV-mediated eGFP labeling with the anterograde tracer cholera toxin subunit B (CTB)24. Mice received an intravitreal injection of CTB and were sacrificed 2 days later for histological analysis. Colocalization of eGFP and CTB signals in optic nerves confirmed the positive tracing of RGC axons (Figure 5A). To further study the changes of signals after axonal injury, mice were subjected to unilateral optic nerve crush (ONC), a model of acute RGC damage25. Briefly, mice were anesthetized, and one optic nerve was exposed and crushed approximately 1 mm behind the globe using fine forceps for 5 s, inducing significant RGC axotomy and subsequent degeneration. Following ONC injury, a marked reduction in both eGFP and CTB intensity was observed, indicating substantial nerve fiber loss and axonal transport disruption. Further analysis of brain sections revealed RGC projections to the Superior Colliculus (SC), a midbrain structure involved in sensorimotor integration and eye movements, and the Lateral Geniculate Nucleus (LGN), the primary relay center for visual information to the visual cortex. After ONC, a decreased GFP intensity was revealed in the contralateral side of the injury (Figure 5B–E). These findings demonstrate the utility of this method for studying RGC axonal degeneration and mapping their projection patterns in vivo, offering a potential alternative to traditional invasive tracing methods.
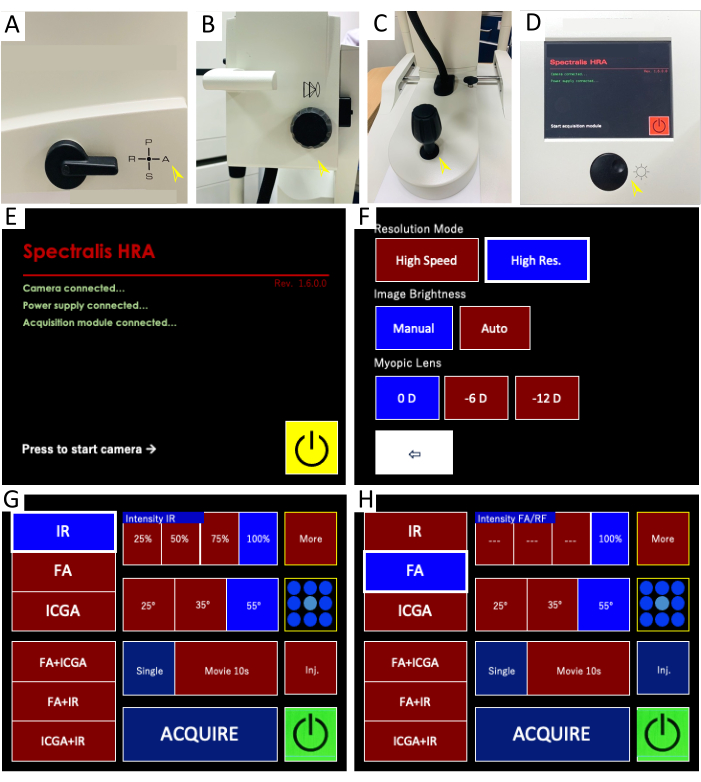
Figure 1: Confocal scanning laser ophthalmoscope (CSLO) imaging system and control panel interface for live retinal imaging. (A–D) Imaging system components: These images depict the key components of the HRA CSLO device. (A) The filter lever is set to position "A" for combined infrared reflectance (IR) and fluorescein angiography (FA) imaging. (B) The focus knob adjusts the fine focus of the objective lens. (C) The micromanipulator allows for precise positioning, alignment, and illumination of the light beam to the retina through multiple degrees of freedom (e.g., X, Y, Z axes) and rotation. (D) The sensitivity knob controls the brightness of both IR and FA images by adjusting the amplification of the detected light signal. Pressing the knob initiates image averaging, which combines multiple frames to reduce noise and enhance the signal-to-noise ratio. (E–H) Control panel interface: The screenshots display the CSLO control panel with the settings used for live image acquisition and visualization. (E) The acquisition control panel is initiated. (F) High-resolution mode is recommended for optimal image quality. (G) The IR imaging settings are highlighted, allowing visualization of retinal structures and blood vessels. (H) The FA imaging settings display options for both single-point and multi-point fluorescence imaging, enabling the detection of fluorescent signals within retinal cells. Please click here to view a larger version of this figure.
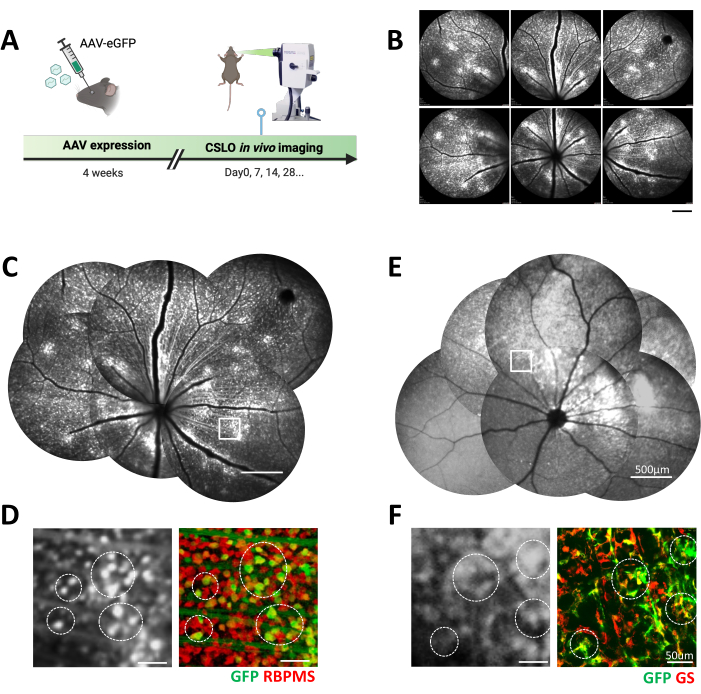
Figure 2: In vivo CSLO imaging of fluorescently labeled retinal cells following AAV transfection in C57 mice. (A) The experimental timeline of CSLO-aided longitudinal imaging after AAV transfection in the mouse eye. (B) Representative CSLO images of the superior and central regions of a C57 mouse retina after 4 weeks of AAV2-hSyn-eGFP expression. Scale bar: 500 µm. (C) A manually stitched retinal montage from (B) showcases the distribution of eGFP-expressing retinal cells. Scale bar: 500 µm. (D) A magnified CSLO image of the boxed area in (C) and the corresponding confocal microscopy image of a retinal flatmount. The colocalization of eGFP (green) with the RGC marker RBPMS (red) confirms AAV-mediated expression in retinal ganglion cells (examples circled). Scale bar: 50 µm. (E) A manually stitched montage of a C57 mouse retina 4 weeks post-injection with AAV9-GfaABC1D-eGFP, demonstrating Müller glia-specific eGFP expression. Scale bar: 500 µm. (F) A magnified CSLO image of the boxed area in (E) and the corresponding confocal microscopy image of a retinal flatmount. The colocalization of eGFP (green) with the Müller glia marker glutamine synthetase (GS, red) confirms AAV-mediated expression in Müller glial cells (examples circled). Scale bar: 50 µm. Please click here to view a larger version of this figure.
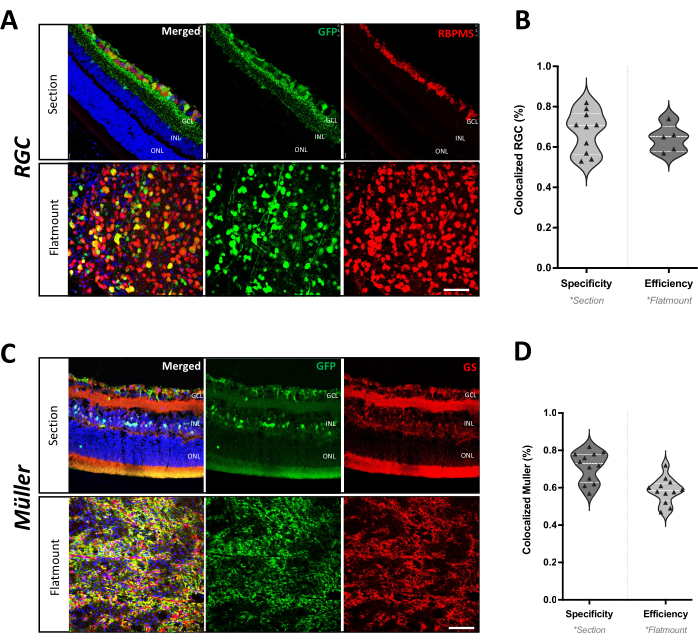
Figure 3: Cell type specificity of AAV-mediated gene delivery in mouse retina. (A) Immunohistochemistry of a retinal section and flatmount from C57 mice after 4 weeks of AAV2-hSyn-eGFP infection. The colocalization of GFP (green) with RBPMS (red) confirms RGC-specific expression. Scale bar: 50 µm. (B) Quantification of AAV2-hSyn-GFP transduction efficiency and specificity in RGCs. Efficiency is represented as the percentage of RBPMS-positive RGCs that also express GFP in the retinal flatmount. Specificity is represented as the percentage of GFP-positive cells that are also positive for RBPMS in the retinal section (n = 6 eyes from 4 mice). (C) Immunofluorescence staining of a retinal section and flatmount from C57 mice after 4 weeks of AAV9-GfaABC1D-eGFP infection. The colocalization of GFP (green) with GS (red) confirms Müller glia-specific expression. Scale bar: 50 µm. (D) Quantification of AAV9-GfaABC1D-GFP transduction efficiency and specificity in Müller glia. Efficiency is represented as the percentage of GS-positive Müller glia that also express GFP in the retinal flatmount. Specificity is represented as the percentage of GFP-positive cells that are also positive for GS in the retinal section (n = 6 eyes from 3 mice). Please click here to view a larger version of this figure.
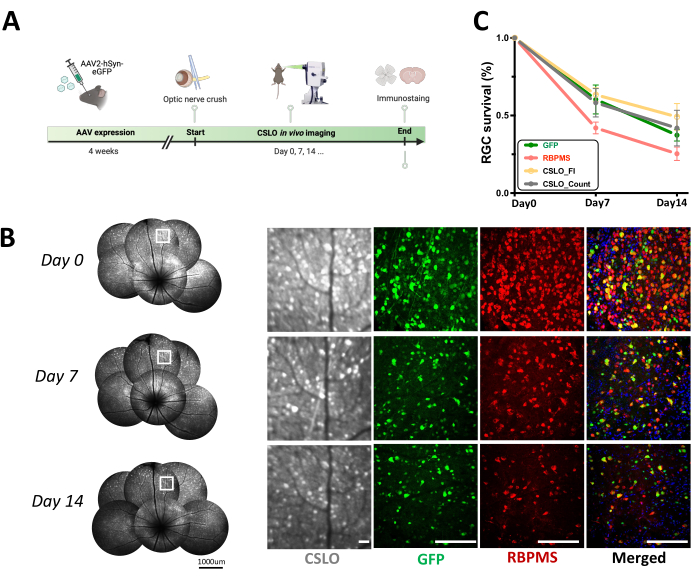
Figure 4: Longitudinal CSLO monitoring of RGC survival following acute optic nerve crush (ONC) in AAV2-hSyn-eGFP mice. (A) A schematic diagram of the experimental timeline of acute optic nerve injury after AAV infection. (B) Longitudinal CSLO imaging of the same retina in an AAV2-hSyn-eGFP mouse at baseline, 7 days, and 14 days post-ONC, visualizing RGCs expressing GFP signals. Magnified images of the boxed areas are shown on the right, alongside representative confocal microscopy images of retinal flatmounts demonstrating RGC morphology and distribution. Scale bars: 100 µm. (C) Quantification of RGC survival over two weeks post-ONC was assessed by measuring the percentage change in both cell number and CSLO fluorescent intensity relative to baseline (mean ± SEM). No significant differences were observed between groups at each time point (one-way ANOVA). Linear regression analysis revealed significant negative correlations between time post-injury and RGC survival metrics based on GFP fluorescence (r² = 0.945, p < 0.001), RBPMS staining (r² = 0.888, p < 0.001), CSLO-cell count (r² = 0.511, p < 0.001), and CSLO fluorescent intensity (r² = 0.7486, p < 0.001). n = 6 mice/group/time point. Please click here to view a larger version of this figure.

Figure 5: AAV-aided mapping of RGC axonal projections along the visual pathway. (A) Representative fluorescence microscopy images of a pair of optic nerves from an AAV2-hSyn-eGFP mouse, demonstrating GFP-labeled RGC axons (green) and colocalization with the anterograde axonal tracer cholera toxin B (CTB, magenta). The upper panel provides an overview of the entire optic nerves (scale bar: 1000 µm), while the lower panel offers magnified views of the boxed regions (scale bar: 100 µm). Two weeks post-ONC injury, both GFP and CTB signals show a marked reduction throughout the optic nerve (left side), indicating RGC axonal loss and disruption along the visual pathway. (B) RGC axonal projections to the superior colliculus (SC). Coronal brain sections from an AAV2-hSyn-eGFP mouse 2 weeks post-ONC highlight GFP-labeled RGC axons (green) and CTB staining (magenta) within the SC. Left (L): contralateral SC; Right (R): ipsilateral SC (ONC-injured side). Scale bar: 1000 µm. (C) A magnified view of the boxed region in (B) demonstrates the colocalization of GFP and CTB signals (merged image), highlighting RGC axonal bundles and their projection patterns within the SC. Scale bar: 100 µm. (D) RGC axonal projections to the lateral geniculate nucleus (LGN) following ONC, revealing GFP-labeled RGC axons (green) and CTB staining (magenta) within the LGN. Left (L): contralateral LGN; Right (R): ipsilateral LGN (ONC-injured side). Scale bar: 1000 µm. (E) A magnified view of the boxed region in (D) highlights RGC axonal bundles and their projection patterns within the LGN. Scale bar: 100 µm. Please click here to view a larger version of this figure.
