By following the steps in Protocols 1 and 2, one should be able to design a simple fly vial rack, and run the model STL file through the slicing program to generate coordinates for the 3D printer. Figure 3A shows a printed unit of the model next to its design. We also hope step 1 can provide the basic skills for one to use the basic shapes available in the Tinkercad platform to create useful apparatuses for the lab. Developing these skills, however, may require constant practice and frequent consultations of the Tinkercad help center45. In Protocol 3, we provide our own designs for apparatuses that we routinely use for general fly maintenance and behavioral studies with adults or larvae. By following Protocol 2, a researcher should be able to use the STL files provided as Supplemental Files 2-6 and 8-10 in Protocol 3 to print tapping mat supports, funnels, Stalkers, Fly Motels, T-Mazes and RING apparatuses. Figure 3B-G shows side-by-side images of the model designs and of the actual printed and assembled apparatuses. It is important to mention that each printer may require minimal adjustments to the printing settings for optimal function; our Supplemental File 1, however, appears to provide a robust initial set of instructions for RepRap 3D printers to print the abovementioned devices.
In addition to providing instructions on how to create and 3D-print apparatuses that can be used for behavioral analyses in Drosophila, we also provide a protocol to perform assays and obtain behavioral measurements using larvae (Protocol 4) and a protocol for high resolution respirometry using larval homogenates (Protocols 5 and 6) to investigate mitochondrial metabolism. The mobility assays provided can easily reveal the impact on larval behavior at cold temperatures, when the fly OXPHOS is directly altered by ubiquitous expression of AOX. AOX-expressing larvae can cover distances ~70% longer than control larvae at the stressful temperature of 12 °C (Figure 4A), apparently due to the ~40% higher number of peristaltic movements of the body wall (Figure 4B). We can also easily associate this change in behavior with changes in mitochondrial function. Figure 5A shows the traces of oxygen consumption as a representative outcome of an experiment using larval homogenates. It is evident that the mitochondria of AOX-expressing larvae have antimycin A-resistant respiration, which is sensitive to the AOX inhibitor propyl-gallate (green arrow in Figure 5A, and Figure 5B), and consistent with previously published data using adult flies and with the expected function of AOX24. Quantification of the different oxygen consumption states also indicates that LN respiration is elevated in AOX-expressing larvae (Figure 5B). This in turn is reflected in the calculations for respiratory control ratio (RCR), which is a parameter that indicates how coupled mitochondria are by dividing the values for OXPHOS respiration by those of Leak respiration46,47. The lower RCR in AOX-expressing larvae suggests mitochondria are less coupled in these flies (Figure 5C). We speculate that the energy that is not being used for ATP synthesis is in fact being dissipated as heat, which gives AOX-expressing larvae more mobility at such an extremely stressful temperature.
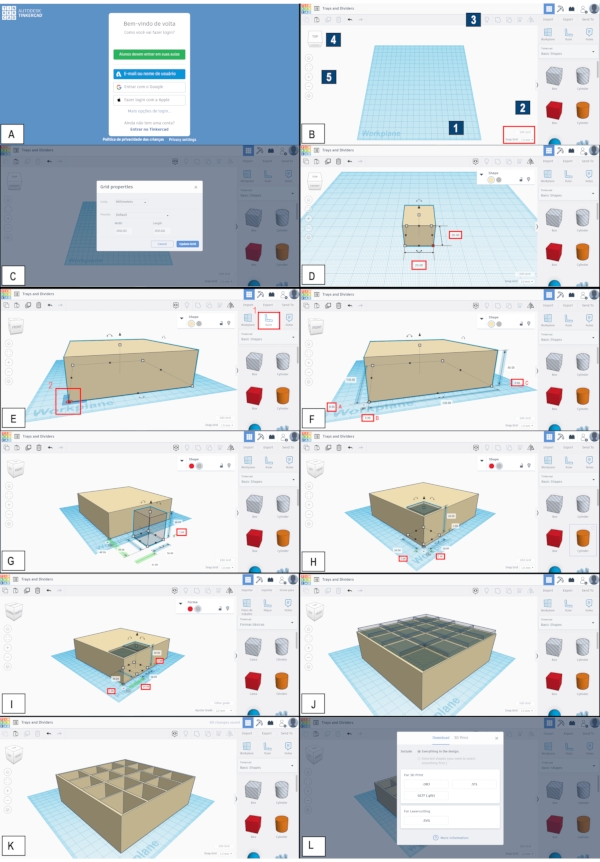
Figure 1. Modeling of a small fly vial rack using Tinkercad. (A) Access to the modeling platform is given online38using the user´s preferred password-protected credentials. (B) General view of the platform. Blue square 1 indicates the workplane, which represents the printer table, where objects are designed. Blue square 2, Basic Shapes menu, where several pre-made shapes are found. Blue square 3, a menu with useful functions, such as mirror, copy, duplicate and combine. Blue square 4, workplane movement menu, which controls rotations of the workplane to better view the object being modeled. Positioning the side of the cube using the mouse´s left button to "Top", "Bottom", "Front", "Back", "Right" and "Left" shows views of the object from different angles. Blue square 5, left menu containing other useful functions, such as zoom in and zoom out, among others. Red square, "Edit Grid" and "Snap Grip" options. (C) The Edit Grid window, where the workplane can be edited. (D) The design of a solid box (future vial rack). Red squares indicate the textboxes in which the dimensions of the box can be typed. (E) The design of a solid box (future vial rack) of 130 (length) x 130 (width) x 40 (height) mm. Red square 1 indicates the tool "Ruler"; red square 2, the lower left vertex of the box, which will become the initial point (x = 0, y = 0, z = 0) of a tridimensional Cartesian coordinate system. (F) The same solid box as in E, with red squares A-C illustrating that the lower left vertex of the box is now 0.00 mm away in all axes from the initial point of a tridimensional Cartesian coordinate system. (G) An empty (hole) box (future space for a fly vial) of 30 x 30 x 40 mm is inserted in the workplane. The red square indicates the textbox in which 2.00 should be typed to elevate the empty box 2 mm from the workplane. (H) The same empty box as in G is positioned inside the initial solid box of 130 x 130 x 40 mm by typing 2.00 in the textboxes indicated with red squares. (I) A second empty box inserted into the design illustrates how the spaces for fly vials are created one by one, based on the first empty box. The red squares indicate the textboxes that must be filled for the proper positioning of the empty boxes. (J) The design is now filled with 16 empty boxes, evenly spaced inside the initial solid box. (K) Final 3D model of the vial rack, which is now a single piece, created by grouping the initial solid box with the 16 empty boxes evenly spaced inside the solid box. See Protocol 1 for details on the whole procedure, including how to save the final model as an STL file (L). Please click here to view a larger version of this figure.
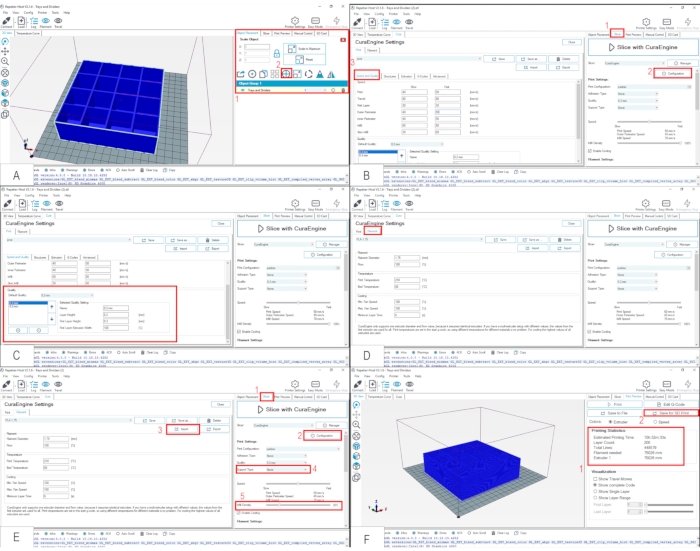
Figure 2. Slicing of the 3D vial rack model and configuring the printer using Repetier-Host. (A) The STL file containing the model of the vial rack (see Protocol 1 and Figure 1 for details) is opened with the Repetier-Host software installed on the computer. The editing menu on the right is indicated by red square 1, and the Center Object button by red square 2. (B) Initial steps to run the slicing software (see Protocol 2 for details). Red square 1 indicates the Slicer tab on the editing menu; red square 2, the Configuration button; red square 3, the Speed and Quality window, where important printer parameters such as velocity, layer thickness and holders can be defined (see the Discussion for details). (C) Detail of the Speed and Quality window. The red square indicates where the thickness of the printing layers (Layer Height and First Layer Height) can be adjusted. (D) In the Filament tab (red square), one is able to check parameters related to the filament to be used, such as the Diameter (which for most filaments is set to 1.75 mm), the Print and Bed Temperatures (respectively, the temperature to melt the filament at the printer extruder, and the temperature of the printer table which helps with adherence), and the Cooling Speed (which in general guarantees that the extruder does not overheat). (E) Final steps to run the slicing software (see Protocol 2 for details). Red squares 1 and 2 same as in (B); red square 3, the Import button, where most of printer´s adjusting parameters can be uploaded from Supplemental File 1; red square 4, Support Type option, which in this particular case of the fly vial rack should be "None"; and red square 5, Infill Density option, which should be 20% for printing most objects. The use of a support, such as for the printing of the T-Maze (see Protocol 3 for details), is necessary when the layers are not to be printed directly onto the printer table or on top of another layer. The support is particularly important to prevent collapse of structural parts that are in an arc format or of closed structures that are hollow inside, for example. (F) Visualization of the printing statistics (red square 1) under the Print Preview tab, which is calculated after the Slice with CuraEngine button (shown in E) is pressed. After pressing the Save for SD Print button (red square 2), the G-Code file should be saved in an SD card to be transported to the printer. Please click here to view a larger version of this figure.
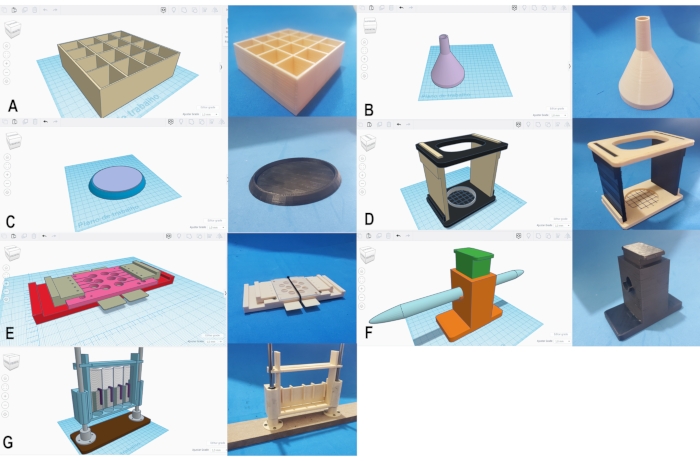
Figure 3. "Homemade" apparatuses for general fly maintenance and behavioral analyses. Side-by-side images of our model designs (left) and of the actual printed apparatuses (right) are shown: small fly rack (A), funnel (B), tapping mat support (C), Stalker (D), Fly Motel (E), T-Maze (F), and RING apparatus (G). Details on how to print and assemble these apparatuses are shown in Protocols 1-3. Note that in F, the picture of the T-Maze does not contain the two 15 mL conical tubes which would give it its "T" shape41. Please click here to view a larger version of this figure.

Figure 4. Higher mobility of AOX-expressing larvae cultured at 12 °C. Measurements of distance crawled (A) and number of body wall contractions (B) per minute by individual larva were obtained as described in Protocol 4. The fly lines used were w1118 (control, AOX-nonexpressor) and 3xtubAOX (AOX-expressor)25. Datapoints indicate means ± standard deviation of 8 biological replicates with 15 technical repetitions each. * indicates significant differences (p<0.01), according to a Student's t-test. Please click here to view a larger version of this figure.
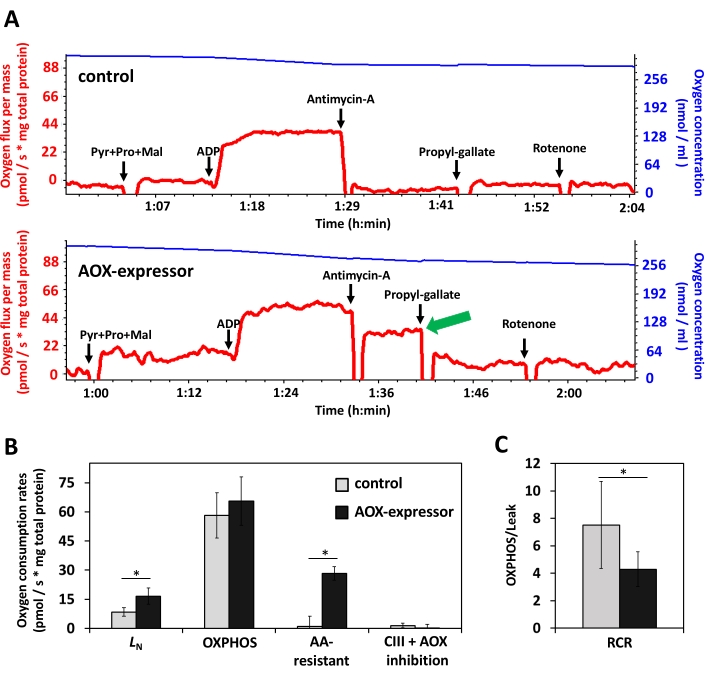
Figure 5. Higher levels of mitochondrial oxygen consumption in AOX-expressing larvae cultured at 12°C. (A) Traces of a representative experiment showing changes in real time oxygen consumption (red line) and oxygen concentration (blue line) in the chambers as the indicated substrates and inhibitors were added (black arrows) to the assay medium containing whole larva homogenates of the w1118 (control, AOX-nonexpressor) and 3xtubAOX (AOX-expressor). Pyr+Pro+Mal indicates addition of pyruvate, proline and malate, which are oxidized inside mitochondria providing NADH, substrate for complex I. The following increase in oxygen consumption is due to Leak respiration without adenylates (LN). Addition of ADP allows the ATP-synthase to release the proton gradient so that the Oxidative Phosphorylative respiratory state (OXPHOS) is achieved. Addition of antimycin A inhibits complex III (CIII), completely abolishing oxygen consumption in the AOX-nonexpressing control sample (top graph), and allowing the antimycin A-resistant respiration activity of AOX (green arrow) to be measured in the AOX-expressing sample (bottom graph). Addition of propyl-gallate, an AOX inhibitor, followed by the complex I inhibitor rotenone, certifies that mitochondrial respiration is totally abolished in both lines, allowing baseline oxygen consumption to be established. The first 50-60 min of the experiments, prior to the addition of Pyr+Pro+Mal, show the traces during stabilization of the oxygraph system, and were deliberately omitted here. The traces before addition of the sample homogenates are used for calculation of air oxygen saturation (see Protocols 5 and 6 for details). (B) Quantification of the oxygen consumption data shown in A, averaged with that of 3-5 other biological replicates (± standard deviation). AA-resistant, antimycin A-resistant respiration; CIII + AOX inhibition, residual respiration. (C) Respiratory control ratio (RCR) was calculated as the average ratio between OXPHOS and LN respirations of the data shown in B and that of 3-5 other biological replicates (+/- standard deviation), to estimate mitochondrial OXPHOS-coupling efficiency. We have previously measured Leak respiration in the presence of oligomycin (LOmy) and determined no differences between LOmy and LN (data not shown). Using LN to compute RCR allows us to measure the antimycin A-resistant respiration by AOX in the same experiment. * indicates statistical differences (p<0.05), according to Student's t-tests. Please click here to view a larger version of this figure.
Supplemental File 1. Please click here to download this File.
Supplemental File 2. Please click here to download this File.
Supplemental File 3. Please click here to download this File.
Supplemental File 4. Please click here to download this File.
Supplemental File 5. Please click here to download this File.
Supplemental File 6. Please click here to download this File.
Supplemental File 7. Please click here to download this File.
Supplemental File 8. Please click here to download this File.
Supplemental File 9. Please click here to download this File.
Supplemental File 10. Please click here to download this File.
Supplemental File 11. Please click here to download this File.
