To analyze the sedimentation profile of Cwc24-TAP and determine whether the Grafix method was effective to stabilize its binding to splicing subcomplexes, we separated total yeast extracts of cells expressing Cwc24-TAP through centrifugation on glycerol gradients, in the presence or absence of glutaraldehyde as a crosslinking agent. Samples of twenty-four 500 μL fractions were then analyzed by slot blot with antibody against the CBP portion of the TAP tag. The results show that in the absence of the crosslinker, Cwc24 is concentrated between fractions 5 and 13 (Figure 1B), corresponding to complexes of about 80 to 200 MDa14. In the presence of the crosslinker, however, the position of Cwc24 on the gradient is shifted to fractions at the bottom of the gradient, corresponding to larger complexes (Figure 1). These results indicate that glutaraldehyde stabilizes the association of Cwc24 with splicing complexes.
In order to establish whether the complexes retaining Cwc24 correspond to the Bact complex, which is formed by snRNPs U2, U5, and U6 and the NTC complex, we compared Cwc24 sedimentation to those of the U5 snRNP subunit Prp8 and the NTC subunit Prp19. Yeast strains expressing either of these proteins fused to the TAP tag were subjected to Grafix, followed by dot blot for the detection of the proteins. These three spliceosome subunits showed similar profiles, sedimenting with larger complexes at the bottom of the gradient. Cwc24 is concentrated in the same fractions, but because it associates transiently with the spliceosome, it is also present in the lighter fractions (Figure 2). Quantification of the dots shows these profiles (Figure 2B).
Interestingly, fractions 11 and 12 are those where Prp8 and Prp19 start to concentrate, suggesting that this is the portion of the gradient where Bact complex starts to sediment. To ascertain that these proteins are bound to complexes in these fractions, aliquots of fractions 11 and 12 were subjected to electrophoresis on native gels and subsequently to western blot. Signals of Prp8 and Prp19 appear as smears that barely enter an 8% acrylamide gel, showing that indeed they are part of large complexes (Figure 3). Cwc24 is also present in fraction 12, but in much lower concentration, consistent with its transient binding to the Bact complex. These results show that glutaraldehyde can be used as a crosslinker to stabilize the binding of transient factors to splicing subcomplexes.
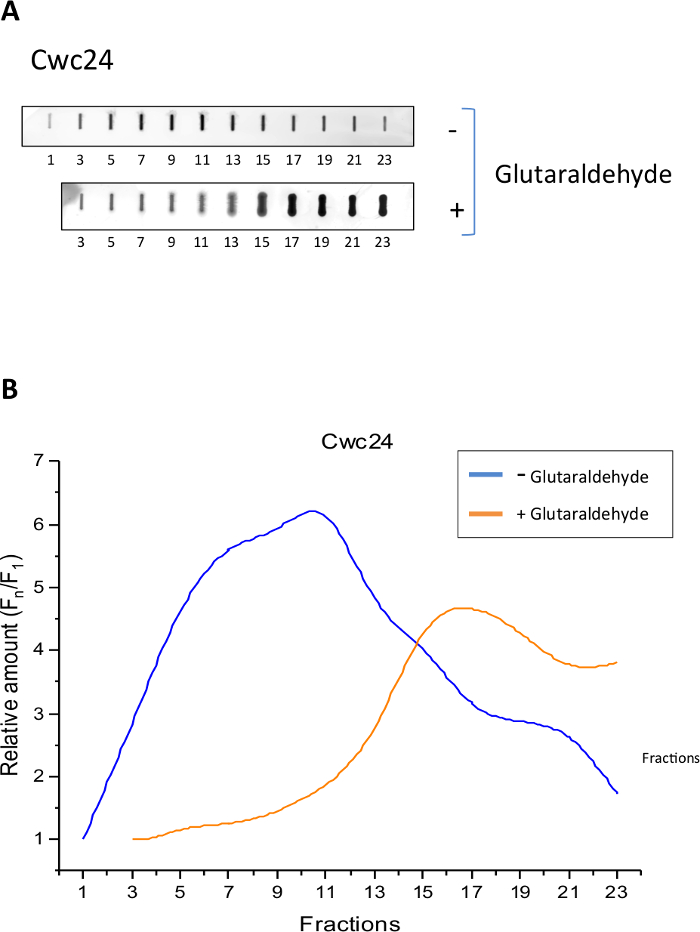
Figure 1: Grafix holds Cwc24 associated to larger complexes. Total yeast extract of cells expressing Cwc24-TAP was separated by centrifugation on glycerol gradients, either in the absence or presence of the crosslinker glutaraldehyde. (A) Odd fractions of the gradient were subjected to slot blot for the immunodetection of Cwc24-TAP with antibody against the CBP portion of the TAP tag. (B) Quantification of Cwc24-TAP using Image J shows the concentration of the protein through the fractions of the glycerol gradient. Presence or absence of glutaraldehyde is indicated. Y axis shows the relative amounts of Cwc24 through the gradient, calculated as the intensity of the signal in the fractions, relative to the first fraction (Fn/F1). Please click here to view a larger version of this figure.
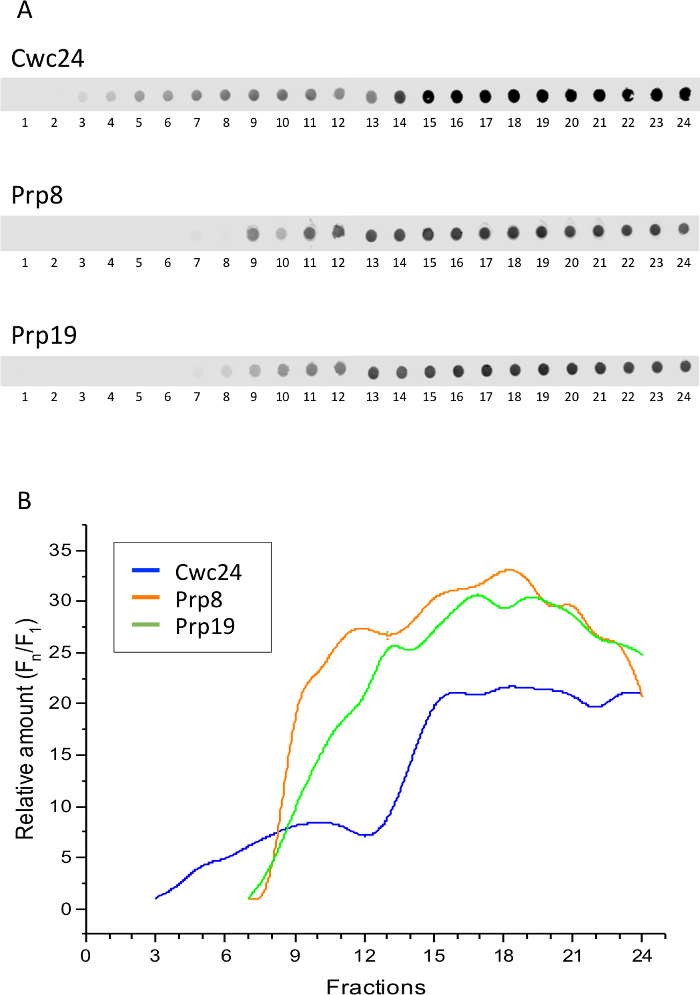
Figure 2: Sedimentation of splicing factors through glycerol/glutaraldehyde gradients. Extracts of yeast cells expressing either Cwc24, Prp8, or Prp19 fused to the TAP tag were loaded on glycerol gradients containing glutaraldehyde and centrifuged for the separation of splicing complexes. (A) Samples of the twenty-four fractions of the gradient were analyzed by dot blot and immunodetection of the TAP-fused proteins with antibody against the CBP portion of the TAP tag. (B) Quantification of the proteins using Image J shows their concentration at the bottom fractions of the glycerol/glutaraldehyde gradient. Y-axis shows the relative amounts of the proteins through the gradient, calculated as the intensity of the signal in the fractions, relative to the first fraction showing a signal above background (Fn/F1). Please click here to view a larger version of this figure.
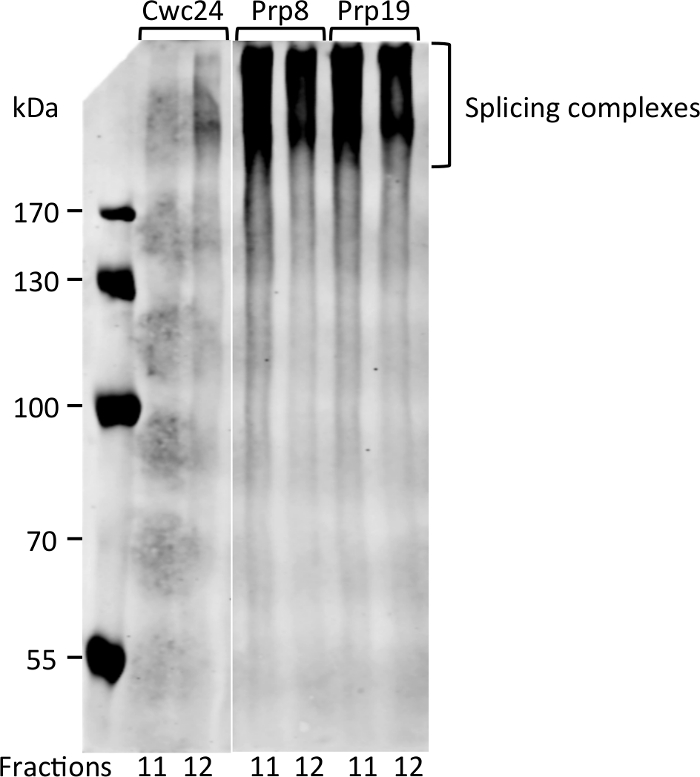
Figure 3: Detection of splicing complexes by native PAGE and immunoblot. Aliquots of fractions 11 and 12 of the gradient shown in Figure 2 were separated by electrophoresis on 8% acrylamide native gel and subjected to immunoblot with antibody against CBP. Proteins are not detected as bands because complexes are too large to separate on native gels. Please click here to view a larger version of this figure.
